Introducción
El sistema nervioso humano percibe, procesa, almacena y ejecuta en respuesta a la información que recibe del medioambiente interno y externo con el fin primordial de asegurar la supervivencia de la especie. Las capacidades claves del cerebro se relacionan a desarrollar habilidades para la supervivencia individual, la selección de pareja y procreación y la protección y cuidados de los dependientes, especialmente los individuos jóvenes. La estrategia primaria para lograr estos objetivos es crear relaciones que nos conectan generando grupos de individuos – familias, clanes, comunidades – que nos han permitido el progreso y el control del medio ambiente. El sistema nervioso humano tiene sistemas neurales complejos y poderosos dedicados a la función de socialización y comunicación, que incluyen desde aspectos motores que permiten la translación, hasta el lenguaje oral, el mejor sistema de comunicación.
El proceso de progresiva adquisición de estas habilidades es lo que denominamos “desarrollo“, consecuencia de procesos cerebrales definidos genéticamente en interacción permanente con el ambiente. Esta relación entre genética o naturaleza versus ambiente o nutrición en sus aspectos más amplios ha generado extensos estudios y discusiones, desde modelos deterministas como el de Gesell y Amartruda (1941), quienes proponen que los factores genéticos y constitucionales ejercen tan primordial influencia sobre el desarrollo, que permiten predecir el futuro de un niño. En contraposición se plantea el Modelo interaccional, que establece que a los factores biológicos implicados en el modelo anterior hay que añadir la interacción del ambiente para predecir el resultado. Así niños con problemas orgánicos constitucionales que crecen en ambientes empobrecidos tendrán mal pronóstico, y aquellos que crecen en ambientes estimulantes, "nutritivos", tendrán un resultado favorable. Más aceptado en la actualidad es el Modelo Transaccional, que subraya la importancia de la plasticidad del niño en desarrollo y de su medio ambiente. El niño, biología y ambiente están continuamente interactuando y provocando cambios el uno en el otro. Por esto la predicción a largo plazo es más difícil que en los modelos anteriores.
Neuro-maduración
El desarrollo del encéfalo humano comienza muy precozmente, alrededor de la 3° a 4° semana de edad gestacional y continúa, aunque a un ritmo decreciente, hasta la adultez. Se ha caracterizado por la ocurrencia de 2 eventos organizacionales mayores. El primer período se inicia con la concepción e incluye los eventos de neurulación, proliferación, migración, y diferenciación. Se ha propuesto que estos eventos son controlados por factores genéticos y epigenéticos (fenómenos no mutacionales pero que varían la expresión de un gen, tales como la metilación y la modificación de histonas) que originan estructuras neurales sensibles a influencias externas. El segundo período es de reorganización, ocurriendo durante la gestación y continuando después del parto. Este estadio se caracteriza por crecimiento dendrítico y axonal, producción de sinapsis, y cambios en la sensibilidad a neurotransmisores.
Neurulación
Se inicia con la formación de la notocorda, que ejerce un papel inductor para la formación de la placa y luego del tubo neural. En los extremos cefálico y caudal queda comunicado con la cavidad amniótica por los neuroporos craneal y caudal. El neuroporo craneal se cierra el día 24-25 y el neuroporo caudal el día 27. La disrupción de este proceso origina la espina o el encéfalo bífido, siendo el cuadro más frecuente el mielomeningocele.
El cierre del neuroporo rostral conduce a la formación del prosencéfalo, cuyo crecimiento desigual genera la formación de un surco entre dos esbozos de vesículas que marca el principio de la diverticulación transversal del prosencéfalo y conduce a la formación del telencéfalo y del diencéfalo.
Diverticulación / clivaje
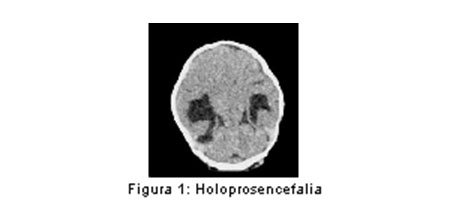
El proceso por el cual el telencéfalo se convierte en una estructura de dos vesículas se denomina diverticulación sagital. La holoprosencefalia (HPE) (figura 1) se debe a la falta de la diverticulación transversal del prosencéfalo. (1,2) La mayoría de los casos de HPE son esporádicos, aislados o asociados a cromosomopatías como la Trisomía 13 (Síndrome de Patau), pero se han descrito formas familiares, generalmente con herencia autosómica dominante con penetrancia reducida y expresión variable. (3,4) Un aspecto interesante se refiere a la descripción de familiares de pacientes con HPE, portadores de la misma mutación, con anomalías craneofaciales menores y neuroimágenes normales, denominadas “microformas HPE”, con riesgo de tener hijos con HPE más graves. Dentro de las anormalidades craneofaciales se describen el hipotelorismo ocular, microcefalia, hipoplasia facial media, anormalidad o ausencia del frenillo labial superior e incisivo maxilar central único. (5)
Poco después de la formación de las vesículas telencefálicas, la mitad ventral de la lámina terminalis telencéfalica aumenta de grosor y se convierte en la placa comisural, que da origen a la comisura anterior, la comisura del hipocampo, el cuerpo calloso y el septum pellucidum. Es probable que la patogénesis de la agenesia del cuerpo calloso sea la ausencia en la placa comisural del área del futuro cuerpo calloso o el fallo de los axones destinados a formar el cuerpo calloso en reconocer esta área. (6) Se ha reconocido la hipoplasia del cuerpo calloso como un marcador de otros trastornos más graves de la organización cerebral. (7)
Estructura interna
Proliferación neuronal “neurogénesis”
Entre el 2º y 4º mes de vida intrauterina se produce la proliferación celular, neurogénesis, en la matriz germinativa periventricular del los ventrículos laterales y 3er ventrículo para las células cerebrales y en el labio rómbico (4º ventrículo) para las del cerebelo. Todas las neuronas se generan en estas regiones, a excepción de las neuronas que originan la capa granulosa externa cerebelosa, que entran en mitosis durante su migración y las del sistema olfativo que mantienen la posibilidad de regeneración por la persistencia de células troncales indiferenciadas en el epitelio olfativo. (8) En los últimos años se ha demostrado neurogénesis en el hipocampo del cerebro adulto de mamíferos, incluyendo humanos, cuyo significado funcional es sujeto de discusión. (9) Se ha reportado que esta neurogénesis puede ser inducida por crisis epilépticas (10) y se ha involucrado en procesos de aprendizaje (11) y en la depresión mayor. (12) Este nuevo conocimiento abre la posibilidad de tratamientos derivados de este proceso. (13) El número de células generadas en el encéfalo fetal es entre un 30 y un 70% superior al número del adulto. Las células “sobrantes” sobreviven por un período de días a semanas, tras lo cual, en forma espontánea, se inicia una cascada de cambios degenerativos y un proceso fisiológico de muerte celular programada o apoptosis.(14) Las injurias que disminuyen la proliferación de neuronas y glía (o aumentan su apoptosis) dan origen a cuadros de microcefalia o micro-lisencefalia; el proceso inverso dará origen a megalencefalias o trastornos más localizados como los hamartomas del complejo esclerosis tuberosa o neoplasias como el gangliocitoma. (15)
Migración neuronal
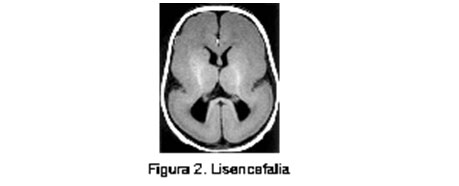
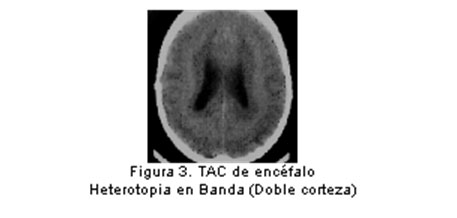
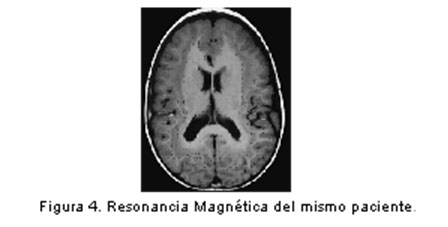
Entre el 3º y 5º mes ocurre la migración de las neuronas, guiadas por procesos gliales en base a señales químicas, como las moléculas de adhesión celular y factores de crecimiento neural. Se han descrito algunos de los eventos moleculares subyacentes al patrón temporo-espacial de desarrollo del sistema nervioso, mediados por genes reguladores que determinan la actividad de otros en una secuencia definida y por lapsos precisos y en regiones específicamente determinadas. (16) Una variedad de insultos intrauterinos y perinatales - como infección, falta de oxígeno, exposición a alcohol o drogas psicotrópicas - pueden alterar la migración de neuronas y la expresión del potencial genético para muchas funciones. Dentro de las malformaciones causadas por anormalidades de la migración neuronal se incluyen el espectro Lisencefalia (figura 2) / Heterotopia Subcortical en Banda (doble corteza), (figura 3 y figura 4) el complejo ”Cobblestone” asociado o no a distrofia muscular congénita y las heterotopias subependimarias (peri ventriculares) o subcorticales. (7)
Organización
Posteriormente toman prioridad la organización y arborización dendrítica, axonogénesis y sinaptogénesis, en la cual las neuronas van estableciendo circuitos neurales de complejidad creciente. Este proceso, además de regulado genéticamente, es sensible a la experiencia, y es la base más importante de la plasticidad del sistema nervioso, (neuroplasticidad) referida a los procesos de desarrollo normal, aprendizaje y recuperación de lesiones. (17) Las conexiones sinápticas pueden ser más rígidas, con un cableado “duro” como en las neuronas que controlan funciones autonómicas - respiración, ritmo cardíaco - en que se requieren conexiones muy precisas, y este proceso está fundamentalmente bajo control genético. Los axones que se originan en la retina están programados para llegar a la corteza visual del cerebro, pero las conexiones locales específicas dependen de señales provenientes del ambiente. En las áreas de asociación, las conexiones son más flexibles, pudiendo ser modificadas hasta la edad adulta.
Desde el punto de vista clínico, la investigación en retardo mental ha demostrado una alteración de la arborización dendrítica como el hallazgo neuropatológico subyacente en la mayoría de los casos de retardo mental, tanto aquellos considerados genéticos como de presumible causa ambiental. (18, 19)
Las malformaciones cerebrales que se relacionan a trastornos de la organización cerebral (incluyendo la migración neuronal tardía) incluyen la polimicrogiria y esquizencefalia. (7)
Mielinización
La mielinización, que significa básicamente el recubrimiento de las conexiones entre las neuronas con una membrana especializada que permite una adecuada transmisión de los impulsos nerviosos, es fundamentalmente un hecho post natal, que ocurre en ciclos, con una secuencia ordenada predeterminada, en dirección caudo - rostral. Al finalizar el 2º trimestre de la gestación se han mielinizado las raíces y médula espinales y se ha iniciado en troncoencéfalo. (14) El haz corticoespinal termina su mielinización a los 2 años, el cuerpo calloso lo hace en la adolescencia y la vía de asociación entre la corteza prefrontal ipsilateral y los lóbulos temporal y parietal lo hace alrededor de los 30 años. (20) La maduración y mielinización normal del encéfalo se relacionan con una reducción de la difusión de agua y aumento de anisotropía de difusión.
Con el desarrollo de nuevas tecnologías de resonancia magnética, se ha logrado identificar alteraciones en la mielina asociadas a retardo del desarrollo. La técnica MRDTI (Magnetic Resonance Difussion Tensor Imaging) detectó diferencias entre los niños con desarrollo normal y aquellos con retraso, los cuales no mostraban alteraciones estructurales en la resonancia magnética convencional. (21)
Neurotransmisores
En el encéfalo maduro, el GABA (ácido gama -aminobutírico) funciona como un neurotransmisor inhibitorio. Se ha descrito que durante el desarrollo del sistema nervioso puede también actuar como factor trófico, influyendo en procesos como proliferación, migración, diferenciación, maduración de la sinapsis y apoptosis, algunos previos a la formación de sinapsis. El GABA media estos procesos a través de la activación de receptores ionotrópicos y metabotrópicos tradicionales, y probablemente por mecanismos sinápticos y no-sinápticos. Sin embargo, las características funcionles del receptor de GABA en el cerebro inmaduro son diferentes e incluso opuestas a lo encontrado en el cerebro del adulto. En el cerebro en desarrollo, el GABA actúa como neurotransmisor excitador, en gran parte debido a una concentración intracelular relativamente alta de cloro en neuronas inmaduras, el cual disminuye durante el desarrollo, permitiendo que GABA llegue a ser progresivamente inhibitorio. (22, 23)
Desarrollo, aprendizaje y neuroplasticidad
Sherrington en 1897 introdujo el término “sinapsis” y que llevó al término plasticidad sináptica para describir los cambios en las propiedades funcionales de una sinapsis como resultado de su actividad. (24) Se ha propuesto que la información puede almacenarse mediante cambios en la eficiencia de la comunicación sináptica. (25)
El aprendizaje se puede describir como el mecanismo a través del cual se adquiere información nueva acerca del ambiente, y la memoria como el mecanismo por el cual ese conocimiento se retiene. Se ha dividido la memoria en aquella explícita, definida como la involucrada en el recuerdo conciente de información que se puede transformar y comunicar en palabras, y la memoria implícita, caracterizada por el recuerdo no conciente de capacidades como habilidades motoras (andar en bicicleta). La memoria explícita depende de la integridad del lóbulo temporal y de estructuras diencefálicas como el hipocampo, subiculum y corteza entorinal. La memoria implícita incluye formas simples de memoria asociativa como condicionamiento clásico, y no asociativas como la habituación, que depende de la integridad del cerebelo y ganglios basales. (26)
Aunque varias estructuras encefálicas juegan un rol en la consolidación de formas de aprendizaje y/o memoria, desde las observaciones realizadas en el paciente H.M., cuyos hipocampos fueron removidos bilateralmente como tratamiento para su epilepsia, se ha reconocido que el hipocampo tiene un rol fundamental en la formación de la memoria declarativa, especialmente para síntesis de la memoria semántica y episódica. (27) Se acepta ampliamente que la formación de la memoria es dependiente de los cambios en eficacia sináptica que permiten el refuerzo de asociaciones entre las neuronas; de hecho, la plasticidad sináptica actividad-dependiente, en las sinapsis apropiadas durante la formación de la memoria, se cree necesaria y suficiente a la vez para el almacenaje de la información.
Ha sido difícil entender el mecanismo por el cual se puede alcanzar la consolidación de conexiones sinápticas, destacando en este esfuerzo el modelo de potenciación a largo plazo, relacionado a activación de receptores NMDA (N-metil-D-aspartato) y receptores metabotrópicos (receptores acoplados a proteínas G las cuales activan o desactivan la síntesis de segundos mensajeros) de glutamato.
Se pueden diferenciar por lo menos dos componentes de memoria: memoria de corto plazo, (de trabajo) de algunas horas de duración, y memoria a largo plazo, que persiste por varios días y a menudo mucho tiempo. En el nivel celular, el almacenaje de la memoria a largo plazo se asocia a la expresión de genes, síntesis de proteínas, y formación de nuevas conexiones sinápticas. (28)
Período vulnerable, periodo crítico y ventanas de oportunidad
La etapa de crecimiento y maduración acelerada del sistema nervioso central (SNC) se considera un “Período Vulnerable” a una serie de noxas, que al actuar van a producir patrones de daño especiales, diferentes a lo que se puede encontrar en un cerebro maduro sometido a las mismas. El resultado puede ser, como en el adulto, la pérdida de funciones adquiridas, pero con mucha mayor frecuencia la no adquisición, retraso, o desviación en la adquisición de habilidades.
El desarrollo secuencial y ordenado del sistema nervioso da origen a otro concepto fundamental, el de “períodos críticos” o, como se prefiere denominar, “períodos sensibles”. Este concepto se refiere a la existencia momentos determinados en la maduración del sistema nervioso en que se establecen las condiciones para lograr una determinada función. Si las estructuras relacionadas a una función se mantienen privadas de las influencias ambientales necesarias para su desarrollo, esta función no se logra en la forma adecuada, incluso si estas influencias logran ejercer su acción en un período posterior. Este conocimiento emergió de estudios clásicos que demostraron que si se tapaba un ojo de un gatito durante sus primeras semanas de vida, se provocaba la pérdida irreversible de la visión de ese ojo debido a la disminución de entradas sinápticas a las neuronas corticales desde el tálamo. (29) Este tema ha recibido gran atención, no sólo de la comunidad científica, sino también de parte de los medios y la comunidad en general, desarrollando el término relacionado de “ventanas de oportunidad”, con importantes implicancias desde el punto de vista educacional a nivel escolar y especialmente preescolar. Un aspecto bien estudiado se relaciona a la adquisición del lenguaje. El aprendizaje de un idioma extranjero como segunda lengua materna es posible sólo hasta antes de la pubertad (período ventana). Posteriormente el aprendizaje es posible, pero con errores gramaticales y dificultad en el acento. Las exploraciones con Tomografía de emisión de positrones (PET) han demostrado que si un niño crece aprendiendo dos idiomas, toda la actividad lingüística se ubica en la misma área del cerebro. Los niños que aprenden un segundo lenguaje más tardíamente muestran dos focos de actividad. (30) En personas de habla inglesa los sonidos r y L se decodifican en partes separadas del cerebro, pero estos sonidos se procesan en la misma parte del cerebro en aquellos en cuya lengua materna (lenguas asiáticas) no distinguen entre estos fonemas. (31)
La posibilidad de inducir un mayor número de conexiones y sinapsis a través de técnicas de estimulación ha sido objeto de un amplio debate. Uno de los que ha recibido mayor atención es un programa de audición de música, específicamente de Mozart, aduciendo efectos positivos en las habilidades cognitivas y, por lo tanto, un mejor desempeño general del individuo frente a múltiples tareas. Una revisión reciente concluye que existe una mejoría específica en el desempeño de habilidades visoespaciales después de escuchar música de Mozart, pero tiene una corta duración y desaparición después de algunos minutos, (10-15 min.), (32) lo que desestimaría el uso comercial que promueve el rápido logro de una inteligencia superior.
Nutrición, desarrollo encefálico y estimulación temprana
Desnutrición calórico –proteica y deficiencias específicas.
La información más relevante sobre la relación entre desnutrición y desarrollo del cerebro se obtuvo de la investigación en ratas de laboratorio. La observación que el retraso del crecimiento del cerebro inducido por malnutrición calórico proteica (MCP) precoz no se recuperaba completamente con la subsecuente alimentación ad libitum (33, 34) dieron credibilidad a la hipótesis que un insulto alimenticio en el período crítico del crecimiento cerebral tiene efectos funcionales permanentes. Sin embargo, esta premisa se ha validado solamente en parte, fundamentalmente por la falta de seguimiento más prolongado. Aunque existe oportunidad para la recuperación, hay también evidencia de lesión cortical permanente. Se ha establecido disminución en la densidad y arborización de dendritas y tamaño de las células corticales. Los sistemas de neurotransmisores también se alteran permanentemente, como el número de receptores del noradrenalina. (32)
La validez de las observaciones en animales de laboratorio y generalización al desarrollo mental de niños desnutridos no ha sido fácil, y se han requerido múltiples estudios longitudinales para acumular evidencia al respecto, sin que todavía se logren respuestas definitivas.
Un estudio en Guatemala demostró que los adolescentes que habían recibido un suplemento de 11,5 gramos de proteína y 682 kilo calorías en período pre y postnatal, durante los primeros 24 meses de la vida lograron mejores resultados en pruebas del conocimiento, cálculo, lectura y vocabulario que aquellos que habían recibido un suplemento menor durante el mismo período, efecto beneficioso restringido a los niños en el nivel socioeconómico más bajo. (35)
Otro estudio se refiere a niños (n = 334) entre 6 y 60 meses provenientes de plantaciones rurales de té en Java, que participaron en un ensayo randomizado de suplemento energético. A los 8 años, sólo los niños que habían recibido el suplemento antes de los 18 meses mostraron mejores resultados que el grupo control, efecto restringido sólo a memoria de trabajo. (36) La suplementación nutricional iniciada a los 18 meses de edad tampoco mostró efectos positivos en un estudio en niños de talla baja en Jamaica, pero agregando estimulación psicomotora a la suplementación nutricional, a través de capacitación a la familia, se observó un aumento estadísticamente significativo en pruebas de inteligencia y de vocabulario respecto al grupo control sin intervención, pero menor comparados a los niños de talla normal. (37) Los niños que rindieron mejor en las pruebas tenían cuidadores con mejor cuociente intelectual verbal y mejor educación, lo que hace difícil distinguir de efectos biológicos relacionados al factor genético en la herencia de la inteligencia.
El acto de alimentar a un niño implica satisfacer su necesidad de nutrientes, pero también tiene una indudable connotación de satisfacción en la esfera emocional. Actualmente es necesario considerar los efectos de la desnutrición como uno de los determinantes del desarrollo, en un marco muy complejo de interacción de factores biológicos, psicológicos y ambientales. Las alteraciones asociadas a desnutrición, como mayor número de hospitalizaciones, falta de actividad y consecuente alteración del vínculo madre – hijo, pueden tener efectos más importantes sobre el desarrollo cognitivo que la falta de nutrientes en sí misma, especialmente si se asocian a situaciones de negligencia o abuso infantil. Los datos disponibles apuntan a que mejores dietas en calidad y cantidad, junto con ambientes estimulantes y la satisfacción de necesidades básicas, podrían prevenir o minimizar los efectos tempranos de la desnutrición en el cerebro. (38)
Algunas deficiencias específicas
Es difícil aislar los efectos de nutrientes específicos debido a las asociaciones e interacciones entre los diversos componentes dietéticos.
Hierro
Una revisión de Cochrane en 2001 se centró en los ensayos placebo-controlados randomizados que examinaban el impacto de la terapia del hierro en el desarrollo en niños menores de 3 años portadores de anemia por deficiencia del hierro. (39) En ensayos de tratamiento a corto plazo con hierro, en 180 niños, no hubo diferencias en funcionamiento mental o motor. Un estudio, encontró, tras 4 meses de tratamiento con hierro, aumentos de 18 puntos en escalas mentales y motoras del Test de Bayley de desarrollo infantil. La mayoría de los investigadores han utilizado grupos control no anémicos, ya que no es ético dejar de tratar la anemia ferropriva. Estudios enfocados a la prevención han arrojado resultados contradictorios. (40)
Zinc
Una revisión de los ensayos publicados sobre los efectos del suplemento de Zinc sobre el comportamiento y desarrollo en edades desde fetal a escolares, mostró que los tres estudios que examinaron actividad reportaron aumento de la actividad relacionada al suplemento de Zinc. De los cinco estudios que examinaron el desarrollo del motor en lactantes y preescolares, tres no mostraron impacto sobre esta área y de cuatro estudios que examinaron el desarrollo mental, tres no encontraron ningún impacto de la suplementación del Zinc. En escolares, dos de tres estudios encontraron un impacto beneficioso en procesos de razonamiento. Se ha planteado que existiría una relación entre Zinc y procesos cognoscitivos en niños más vulnerables, pero se destaca la necesidad de investigación adicional en este aspecto. (41)
Ácidos grasos poliinsaturados de cadena larga y lactancia materna
Existe consenso sobre las indiscutibles ventajas de la lactancia natural en una variedad de aspectos del desarrollo del niño. Un aspecto estudiado es el aporte de ácidos grasos poliinsaturados. Se ha demostrado que los ácidos grasos poliinsaturados de cadena muy larga presentes en la leche materna se asocian a una mejor maduración de la agudeza visual. (42) Sin embargo, algunos de los beneficios en relación al desarrollo de la inteligencia requieren más estudios. En una revisión reciente, se identificaron 40 publicaciones pertinentes (1929 a 2001), de las cuales el 68% concluía que la lactancia natural promueve la inteligencia, pero se consideró que muchos tenían defectos metodológicos, y que la evidencia derivada de los dos estudios de alta más calidad (43, 44) es menos convincente respecto a este punto. (45)
Neurobiología del desarrollo motor
El desarrollo de la motricidad gruesa no requiere mayor estimulación del ambiente, por lo que su retraso se debe habitualmente a causas biológicas. Existe variación normal en la adquisición de los hitos del desarrollo, pero en el desarrollo motor grueso, como en la adquisición de la marcha, esta variación es menor que en otras áreas. Un estudio en 404 niños con retraso de la marcha mostró que un tercio de los niños que a los 18 meses no ha logrado dar 5 pasos en forma independiente, presentarán alguna patología. (46) En los niños con retraso de la marcha y antecedentes de haber sido recién nacidos pequeños para la edad gestacional o de haber requerido admisión a sala de cuidados especiales, la probabilidad aumenta al 56%, con la mitad de los casos diagnosticados como parálisis cerebral. (47) La displasia de caderas no retrasa la adquisición de hitos motores. (48)
Otras causas de retardo motor incluyen la parálisis cerebral en cualquiera de sus formas, las enfermedades neuromusculares, hidrocefalia, tumores -especialmente cerebelosos que alteran el desarrollo del equilibrio- y enfermedades degenerativas /metabólicas, como la ataxia - telangectasia o la leucodistrofia metacromática entre muchas otras.
Un desarrollo motor grueso dentro de lo esperado no es garantía de inteligencia normal a futuro. Los niños con retardo mental (RM), sin otras discapacidades, en general adquieren la marcha independiente a edades más tardías que los niños con inteligencia normal, pero dentro de cada nivel de RM existen niños que caminan a edades comparables a los normales. (49) Se ha reportado que sólo el 62,2 % de los niños con RM grave y 38 % de los con deficiencia moderada caminan después de los 12 meses, lo que demuestra que el desarrollo motor puede ser aparentemente normal el primer año de vida. (50)
Mucho más predictivo es el uso que hace el niño de sus manos en relación a la exploración del ambiente. La desaparición de los reflejos primitivos y la maduración de la función visual permiten al lactante el uso de sus manos, y alrededor de los tres meses, cuando lo cercano puede ser enfocado y logra recibir información simultánea de la vista y el tacto, se establece la base de las futuras habilidades visomotoras. Desde los 3 a los 6 meses progresivamente logra la prensión voluntaria y visualmente guiada, primero en el plano lateral y luego en la línea media. La dominancia (dexteridad o zurdera) no se desarrolla hasta después del primer año y está claramente definida sólo después de los 2 años. Un lactante que muestra una clara predilección por una de sus manos y cruza la línea media para tomar un objeto debe hacer pensar en una lesión motora como una hemiparesia.
La manipulación de objetos refleja la progresiva comprensión del mundo que lo rodea. A los 9 meses el niño examina los objetos en forma sistemática, gracias a la capacidad de procesar información en forma simultánea y no secuencial como anteriormente. Manifestaciones importantes de este desarrollo cognitivo son la aparición alrededor de los 9 meses del sentido de permanencia de los objetos que demuestra la representación simbólica de los objetos y la relación de causalidad.
Desarrollo de la comunicación y lenguaje
En esta área es donde el debate sobre la importancia relativa de mecanismos biológicos y ambientales en su desarrollo ha recibido mayor atención. La pregunta es en que medida habilidades cognitivas como el lenguaje son el resultado de estructuras y predisposiciones específicas y genéticamente codificadas.
Se ha planteado que si los cerebros de los niños están predispuestos en forma innata a aprender lenguaje, con la exposición adecuada todos los niños con cerebros normales deben, sin instrucción, aprender la lengua de una manera relativamente uniforme. Si esta hipótesis es correcta, la capacidad de adquirir lenguaje debe ser autónoma tanto funcional como anatómica de otras capacidades, y las lesiones del desarrollo o adquiridas pueden deteriorar o por el contrario preservar específicamente la capacidad de aprender lenguaje. Si se acepta la ponencia que la capacidad de aprendizaje del lenguaje no es innata, la instrucción debe ser necesaria para aprenderlo, el curso de la adquisición debiera variar considerablemente en cada persona (quizás en función de la calidad de la instrucción), no debiera haber período crítico para la adquisición, y no debe haber evidencia para la especificidad funcional o anatómica del lenguaje. (51)
Revisiones de estudios de familias, y meta-análisis han revelado que los factores hereditarios cumplen un importante rol en los trastornos específicos del lenguaje, en la dislexia y en el desarrollo del lenguaje escrito y hablado normal. (52)
El trastorno específico del lenguaje (TEL) es un trastorno del desarrollo definido clínicamente como incapacidad de adquirir o usar el lenguaje en forma adecuada, existiendo una educación formal y un ambiente favorable, y en ausencia de retraso mental, déficit motor o sensorial. El TEL afecta aproximadamente al 7% de los niños que entran a jardín infantil (53) y se asocia generalmente a resultados académicos pobres si no se resuelve. (54, 55)
Se acepta que la mayoría de las dislexias se relacionan a dificultades en el procesamiento del lenguaje, primariamente a nivel del procesamiento fonológico de los sonidos del habla, específicamente percepción o conciencia fonológica, que es la habilidad de reconocer y manipular la estructura de sonidos de las palabras. (56, 57)
Se ha descrito alteración de la actividad cerebral durante la lectura en niños y adultos disléxicos, (58, 59) sin embargo se ha reportado que intervenciones terapéuticas mediante programas computacionales serían capaces de generar cambios en la función cerebral, evidenciados a través de resonancia magnética funcional (FRM), con mejoría clínica en lenguaje, lectura y procesamiento fonológico. (60)
Al evaluar un niño es necesario diferenciar el habla, definida como el acto motor de comunicarse mediante la articulación de expresión verbal, del lenguaje, entendido como el conocimiento y uso de un sistema de símbolos usado para la comunicación interpersonal. El desarrollo del lenguaje se considera un muy sensible indicador del estado de desarrollo y es un factor predictivo de trastornos posteriores. El lenguaje comprende no sólo el habla, sino también el escuchar, comprender, y la comunicación por medios no verbales.
Etiología de retraso en adquisición de lenguaje
Aunque retraso del lenguaje no es sinónimo de retraso mental, éste es la causa más frecuente de retraso del lenguaje, y todos los niños con RM presentan algún grado de retraso del lenguaje. (61)
Aunque menos frecuente que el retardo mental, más importante aun es la pesquisa de hipoacusia. Entre 1 a 6 por mil recién nacidos presenta pérdida auditiva significativa, (62) la que es necesario detectar en forma precoz (al menos antes de los 3 meses de edad) para lograr un mejor desarrollo especialmente de lenguaje. (63)
Un aspecto debatido es el rol de la otitis media con líquido como causa de retraso de lenguaje. Una revisión reciente (meta-análisis) mostró una asociación de leve a inexistente entre la hipoacusia asociada a esta entidad y el posterior desarrollo de habla y lenguaje. (64)
Otros cuadros a considerar en un niño con retardo en la adquisición de las habilidades lingüísticas incluyen el autismo infantil, los trastornos específicos del lenguaje, algunas enfermedades metabólicas, trastornos motores severos (anartria) y un ambiente extremadamente empobrecido.
Repercusiones de los trastornos de lenguaje
Un estudio en Nueva Zelanda identificó niños de tres años con retraso de lenguaje. Un 2.6 % presentaban compromiso sólo de la comprensión, y porcentajes similares presentaban déficit de la expresión y globales. Evaluados a los 7, 9 y 11 años, comparados con los controles, presentaban en promedio resultados estadísticamente significativos más bajos en cuociente de inteligencia y pruebas de lectura. Los grupos con retraso de la comprensión y retraso global tenían considerablemente más problemas del comportamiento que el resto de la muestra. Los resultados de este estudio confirman la importancia de las alteraciones del lenguaje como predictores de inteligencia bajo el promedio, problemas de la lectura y del comportamiento. (65) Esto hace tener especial cuidado con el “retraso simple de lenguaje”, diagnóstico frecuente especialmente entre profesionales de la salud no médicos, considerado de buen pronóstico pese a la escasa información disponible, (66) y que es un término que no aparece en las clasificaciones de trastornos del lenguaje más aceptadas. (67)
El rol de la terapia de lenguaje también ha sido objeto de estudio, buscando la mejor manera de ayudar a este grupo de niños. En una revisión reciente, se evidencia la falta de estudios metodológicamente adecuados y se concluye que la terapia de lenguaje sería efectiva, comparada con un control expectante, sólo en aquellos niños sin trastornos de la comprensión. (68)
Conclusión
Con el conocimiento adecuado y una visión crítica sobre las intervenciones disponibles, el pediatra, al incluir dentro de los controles periódicos de niño sano la evaluación del desarrollo psicomotor, tiene la capacidad para tomar las medidas necesarias para su diagnóstico adecuado en el caso de detectar una alteración y guiar a la familia en la búsqueda de los apoyos necesarios y elección de las intervenciones más adecuadas a cada niño.
Referencias
- Volpe J. Neurology of the newborn. 3 ed. Philadelphia: WB Saunders Co; 1995.
- Leech RW, Shuman RM. Holoprosencephaly and related midline cerebral anomalies: A review. J Child Neurol 1986; 1: 318.
- Olsen C., Hughes J., Youngblood L., Sharpe-Stimac M. The epidemiology of holoprosencephaly and phenotypic characteristics of affected children: New York State, 1984–1989. Am J Med Genet 1997; 73: 217–26.
- Muenke M., Gurrieri F., Bay C. et al. Linkage of a human malformation, familial holoprosencephaly, to chromosome 7 and evidence for genetic heterogeneity. Proc Natl Acad Sci USA1994; 91: 8102–6. [Citado 14 abril 2005] Disponible en Internet: http://www.pnas.org/cgi/content/abstract/91/17/8102
- Heussler H., Suri M., Young I., Muenke M. Extreme variability of expression of a Sonic Hedgehog mutation: attention difficulties and holoprosencephaly. Arch Dis Child. 2002 Apr; 86 (4): 293-6.
- Gelot A, Esperandieu O, Pompidou A. Histogenesis of the corpus callosum. Neurochirurgie. 1998; 44 (suppl 1): 6173.
- Schaefer G., Bodensteiner J., Thompson J. Jr, Wilson D. Clinical and morphometric analysis of the hypoplastic corpus callosum. Arch Neurol. 1991 Sep; 48 (9): 933-6.
- Barkovich A., Gressens P., Evrad P. Formation, maturation and disorders of brain neocortex. AJNR. 1992; 13: 423- 446
- Kempermann G., Wiskott .L, Gage F. Functional significance of adult neurogenesis. Curr Opin Neurobiol. 2004 Apr; 14(2): 186-91.
- Parent J. The role of seizure-induced neurogenesis in epileptogenesis and brain repair, Epilepsy Research. 50 (1-2) (2002) pp. 179 - 189.
- T.J. Shors G., Miesegaes A., Beylin M., Zhao T., Rydel and E. Gould. Neurogenesis in the adult is involved in the formation of trace memories. Nature 410 (2001), pp. 372–376. [Citado 14 abril 2005] Disponible en Internet:
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?holding=npg&cmd=Retrieve&db=PubMed&list_uids=11268214&dopt=Abstract
- Santarelli L., Saxe M., Gross C., Surget A., Battaglia F., Dulawa S., Weisstaub N., Lee J., Duman R., Arancio O. et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301 (2003), pp. 805–809
- Crespel A., Baldy-Moulinier M., Lerner Natoli M. Neurogenesis in the adult brain: the demise of a dogma and the advent of new treatments. Rev Neurol (Paris). 2004; 160 (12): 1150-8.
- Ferrer I., Soriano E., Del Río J. Cell death and removal in the cerebral cortex during development. Progr Neurobiol 1992; 39: 1-43
- Barkovich A., Kuzniecky R., Jackson G., Guerrini R., Dobyns W. Classification system for malformations of cortical development Neurology 2001; 57: 2168-2178
- Holland P. Homebox genes and the vertebrate head Development 1988; 103: S17- S24
- Lenn N. Brain plasticity and regeneration. AJNR.1992; 13: 505-515
- Huttenlocher P. Dendritic and synaptic pathology in mental retardation. Pediat Neurol1991; 7: 79–85
- Kaufmann W. Cytoskeletal determinants of dendritic development and function: implications for mental retardation. Devl Neuropsychol 1999; 16: 341–346
- Yakovlev P., Lecours A-R. The myelination cycles of regional maturation of the brain. In Minkowsky A. (ed): Regional Development of the brain in early life. Philadelphia, Davis, 1967: 3-70
- Filippi C., Lin D., Tsiouris A., Watts R., Packard A., Heier L., Ulug A. Diffusion-tensor MR imaging in children with developmental delay: preliminary findings. Radiology. 2003 Oct; 229(1): 44-50
- Owens D., Kriegstein A. Is there more to GABA than synaptic inhibition? Nat Rev Neurosci. 2002 Sep; 3(9): 715-27.
- Lujan R., Shigemoto R., Lopez-Bendito G. Glutamate and GABA receptor signaling in the developing brain. Neuroscience. 2005; 130(3): 567-80.
- Bennett M. The early history of the synapse: from Plato to Sherrington. Brain Res Bull. 1999 Sep 15; 50(2): 95-118
- Bliss T., Collingridge G. A synaptic model of memory: long-term potentiation in the hippocampus. Nature 1993; 361: 31-9.
- Squire L. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol Rev 99: 195–231, 1992.
- Scoville R., Milner B. Loss of recent memory after bilateral hippocampal lesions. J Neurol Neurosurg Psychiatry 20: 11–21, 1957
- Lynch M. Long-Term Potentiation and Memory Physiol. Rev. 84: 87-136, 2004
- Hubel D., Wiesel T. The period of susceptibility to the physiological effects of unilateral eye closure in kittens. J Physiol 1970; 206: 419
- Kim K., Relkin N., Lee K. et al. Distinct cortical areas associated with native and second languages. Nature 1997; 388: 171–4.
- Chugani H., Phelps M., Mazziotta J. PET study of human brain functional development. Ann Neurol 1987; 22: 487–97.
- Rauscher F., Shaw G., Ky K. Listening to Mozart enhances spatial-temporal reasoning: towards a neurophysiological basis. Neurosci Lett. 1995 Feb 6; 185(1): 44-7
- Dobbing J., Smart J. Vulnerability of developing brain and behaviour. Br Med Bull. 1974 May;30 (2): 164-8
- Levitsky D. & Strupp B. (1995) Malnutrition and the brain, changing concepts, changing concerns. J Nutr. 125 (suppl.): 2212S–2220S.
- Pollitt E., Gorman K., Engle P., Martonell R., Rivera J. Early supplementary feeding and cognition. Monogr. Soc. Res. Child Dev. 1993; 58: 7
- Pollitt E., Watkins, W., Husaini M. Three-month nutritional supplementation in Indonesian infants and toddlers benefits memory function 8 y later. Am. J. Clin. Nutr. 1997; 66: 1357–1363.
- Walker S., Grantham-McGregor S., Powell C., Chang S. Effects of growth restriction in early childhood on growth, IQ and cognition at age 11 to 12 years and the benefits of nutritional supplementation and psychosocial stimulation. J Pediatr 2000; 137: 36-41
- Pollitt E. Developmental Sequel from Early Nutritional Deficiencies: Conclusive and Probability Judgments. J. Nutr. 130: 350S–353S, 2000. [Citado 14 abril 2005] Disponible en internet: http://www.nutrition.org/cgi/content/full/130/2/350S
- Martins S., Logan S. & Gilbert R. (2001) Iron therapy for improving psychomotor development and cognitive function in children under the age of three with iron deficiency anaemia (Cochrane Review). In: The Cochrane Library, Issue 4. Update Software, Oxford, UK
- Black M. Micronutrient deficiencies and cognitive functioning. BJNutr. 2003 Nov; 133(11 Suppl 2): 3927S-3931S
- Black M. The evidence linking zinc deficiency with children's cognitive and motor functioning. J Nutr. 2003 May; 133 (5 Suppl 1): 1473S-6S.
- Uauy R., Hoffman D., Mena P., Llanos A., Birch E. Term infant studies of DHA and ARA supplementation on neurodevelopment: results of randomized controlled trials. J Pediatr. 2003 Oct; 143 (4 Suppl): S17-25
- Johnson D., Swank P., Howie V., Baldwin C., Owen M. Breast feeding and children’s intelligence. Psychol Rep. 1996; 79: 1179–1185
- Wigg N., Tong S., McMichael A., Baghurst P., Vimpani G., Roberts R. Does breastfeeding at six months predict cognitive development? Aust N Z J Public Health. 1998; 22: 232–236
- Jain A., Concato J., Leventhal J. How good is the evidence linking breastfeeding and intelligence? Pediatrics. 2002 Jun; 109(6): 1044-53
- Chaplais J DE Z, MacFarlane A. A review of 404 late walkers. Arch Dis Child 1984; 59: 512-6.
- Johnson A., Goddard O., Ashurst H. Is late walking a marker of morbidity? Steering Committee, Oxford Region Child Development Project. Arch Dis Child. 1990; 65(5): 486-8.
- Kamath S., Bennet G. Does developmental dysplasia of the hip cause a delay in walking? J Pediatr Orthop. 2004 May-Jun; 24(3): 265
- Hreidarsson S., Shapiro B., Capute A. Age of walking in the cognitively impaired.Clin Pediatr (Phila). 1983 Apr; 22(4): 248-50.
- von Wendt L., Makinen H., Rantakallio P. Psychomotor development in the first year and mental retardation--a prospective study.J Ment Defic Res. 1984 Sep; 28 (Pt 3):219-25.
- Stromswold, K. (1995) The cognitive and neural bases of language acquisition. The cognitive neurosciences, ed M. Gazzaniga. Cambridge, MA: MIT Press.
[Citado 14 abril 2005] Disponible en Internet:
http://ruccs.rutgers.edu/forums/Intro_to_Cognitive_Science/Stromswold_Cognitive_and_Neural_Aspects.pdf
- Stromswold K. 2001. The heritability of language: A review and metaanalysis of twin, adoption and linkage studies. Language 77, 647-723. [Citado 14 abril 2005] Disponible en Internet: http://ruccs.rutgers.edu/~karin/stromswoldLANG.pdf
- Tomblin JB, Records NL, Buckwalter P, Zhang X, Smith E, O’Brien M: Prevalence of specific language impairment in kindergarten children. J Speech Lang Hear Res 1997; 40: 1245–1260.
- Stothard S., Snowling J. Bishop D., Chipchase B., Kaplan C. Language-impaired preschoolers: a follow-up into adolescence. J Speech Lang Hear Res 1998; 41: 407–418.
- Snowling M., Adams J., Bishop D., Stothard S. Educational attainments of school leavers with a preschool history of speech-language impairments. Int J Lang Commun Dis 2001; 36: 173–183.
- Wagner R., Torgenson, J. The nature of phonological processing and its causal role in the acquisition of reading skills. Psychol. Bull. (1987) 101, 192-12
- Wagner R., Torgesen J., Rashotte C., Hecht, S., Barker T., Burgess S., Donahue J., Garon T. Changing relations between phonological processing abilities and word-level reading as children develop from beginning to skilled readers: A 5-year longitudinal study. Develop. Psychol. (1997) 33: 468-479.
- Temple E., Poldrack R., Salidis J., Deutsch, G., Tallal, P. , Merzenich M. & Gabrieli J. Disrupted neural responses to phonological and orthographic processing in dyslexic children: an fMRI study. NeuroReport (2001) 12: 299-307
- Temple E. Brain mechanisms in normal and dyslexic readers. Curr. Opin. Neurobiol. (2002) 12: 178-183.
- Temple E., Deutsch G., Poldrack R., Miller S., Tallal P., Merzenich M., Gabriela J. Neural deficits in children with dyslexia ameliorated by behavioral remediation: evidence from functional MRI. Proc Natl Acad Sci U S A. 2003 4; 100(5): 2860-5. [citado 16 marzo 2005] Disponible en Internet: http://www.pnas.org/cgi/content/full/100/5/2860
- Coplan J. Evaluation of the child with delayed speech or language. Pediatr Ann 1985; 14: 203-8.
- Bachmann K., Arvedson J. Early identification and intervention for children who are hearing impaired. Pediatr Rev.1998; 19: 155 –165
- Yoshinaga-Itano C., Sedey AL, Coulter B., Mehl A. Language of early- and later-identified children with hearing loss. Pediatrics.1998; 102: 1168 –117.
- Roberts J., Rosenfeld R., Zeisel S. Otitis media and speech and language: a meta-analysis of prospective studies. Pediatrics. 2004 Mar;113: e238-48. [Citado 14 abril 2005] Disponible en Internet: http://pediatrics.aappublications.org/cgi/content/full/113/3/e238
- Silva P., Williams S., Mcgee R. A longitudinal study of children with developmental language delay at age three: later intelligence, reading and behaviour problems. Dev Med Child Neurol 1987; 29: 63040.
- McRae K., Vickar E. Simple developmental speech delay: a follow-up study. Dev Med Child Neurol 1991; 33: 868-74.
- Rapin I., Allen D. Syndromes of developmental dysphasia and adult aphasia. In Plum F, ed. Language, communication and the brain. New York: Raven Press; 1988. p. 57-75.
- Law J., Garrett Z., Nye C. The efficacy of treatment for children with developmental speech and language delay/disorder:meta-analysis. J Speech Lang Hear Res. 2004 Aug; 47(4): 924-43. [Citado 14 abril 2005] Disponible en internet: http://www.findarticles.com/p/articles/mi_qa3788/is_200408/ai_n9444374
|